Untitled 59
Jumping Genes: Transposable Genetic Elements The term transposable
genetic element is
the most generic term used to describe a genetic element that can occasionally
move (transpose)
from one position on a chromosome to another position
- on the same chromosome or
- on a different chromosome.
Specific types of these elements have other names, including
- jumping genes
- roving genes
- controlling elements
- cassettes
- transposons
Such elements often cause abnormalities in gene function at the loci where they
insert--most often by disrupting
normal expression of the gene.
Tranposable Genetic Elements were first discovered and described in corn by Barbara
McClintock, who won the Nobel Prize in Physiology or Medicine in 1983 for
her lifelong work.
Transposable elements were not isolated at the molecular level until they were
studied in yeast and Drosophila.
Insertion sequences, transposons and phage
µ are some of the transposable
elements found in bacteria. These TGEs work
at the level of the DNA molecule.
In eukaryotic cells, transposable elements have been found in corn (maize),
yeast, Drosophila and
some mammalian systems. In eukaryotes, transposable elements can be responsible
for rearrangements of entire chromosomes by causing breakages. In some cases,
and RNA intermediate is utilized during transposition.
Discovery of Tranposable Elements in Corn (Zea mays) In 1938, Marcus
Rhoades reported unexpected (non-Mendelian) ratios in certain corn crosses.
This would mean that the presence of an Dt allele allowed spots of pigment to
form in a corn kernel that was genetically supposed to be colorless. Not very
parsimonious.A second hypothesis was proposed:
The original parent plant must have been A1 A1 dt
dt, and the plant resulting from the mutations was A1 a1 Dt
dt.This still didn't explain the dotted kernels. Could the dots have
resulted from somatic mutations? Maybe...but there would have had to be a
tremendous number of separate somatic mutations to account for all those
dots.
A lucky break...
Rhoades found a male corn plant in which the anthers exhibited the dotted
pigment pattern. He used pollen from these to test cross with a1a1 females.
Some of the progeny were completely pigmented. This suggested that something in
the dotted individuals' genes could somehow "reawaken" the ability to produce
pigment in the dotted individuals offspring--but not always. What was going on?
The a1 a1 allele
is the first known example of an unstable
mutant allele--one in which reverse mutations occur at a very high rate.
The Ds element In the 1940s, Barbara McClintock noted in her cytological
studies of corn chromosomes that in one strain of corn, chromosome 9 readly
broke at a specific site. She hypothesized that the break was due to the
presence of two genetic factors she named >Ds (for
"Dissociation"--this one was located at the breakage site) and Ac (for
"Activator"--because the Ds site would not break unless Ac was present).
But when she tried to map them...they wouldn't hold still! From this, she
predicted that the two elements were mobile,
and could actually change places within the genome and
She also found rare, unusual and unexpected corn kernel phenotypes in the
offspring of her corn crosses:

In this example, the presence of Ac causes Ds to break, and the acentric
fragment is lost. The result is hemizygosity at all the loci carried away on the
lost fragment, allowing recessive phenotypes to be expressed in the cells
derived from the single cell in which this breakage occurred early in the corn
kernel's development.
 In this example, Ds in inserted into C early in the kernel's development,
suppressing pigment production. In a few progenitor cells, Ds later pops out.
This allows the normal function of C to resume, and the areas where Ds has
excised are now able to produce pigment. This is an example of how a
transposable element (Ds) can produce an unstable
phenotype: expression changes in different cell lines and at different times
because you never know when Ds is going to pop out of the gene and allow it to
resume function.
In this example, Ds in inserted into C early in the kernel's development,
suppressing pigment production. In a few progenitor cells, Ds later pops out.
This allows the normal function of C to resume, and the areas where Ds has
excised are now able to produce pigment. This is an example of how a
transposable element (Ds) can produce an unstable
phenotype: expression changes in different cell lines and at different times
because you never know when Ds is going to pop out of the gene and allow it to
resume function.Autonomous and Nonautonomous Elements
In plants, there are two types of transposable elements:
1. autonomous elements (these
can tranpose by themselves)2. nonautonomous
elements (which need the input
(i.e., enzymatic product) of a separate element in order to transpose).
Insertion of either type of element into a gene causes that gene to be
disrupted, producing a mutant phenotype.In Rhoades' early study, Dt was that
separate element: it supplied the factors promoting the transposition of a gene
segment, and insertion of that segment into the pigment gene (A) disrupted the
wild type allele's (A1) function, causing the mutant, unpigmented a1 phenotype.
Insertion of an autonomous element is unstable, because it can direct its own
transposition over and over. The mutation can occur in each generation; the
allele produced by the insertion is called a mutable
allele because of its instability.
Insertion of a nonautonomous element is stable, because it needs the products
of the autonomous element in order to transpose and produce the mutant allele.
Let's look:
 Top row: Wild type pigmented kernel.
Second row: Ds is inserted into pigment gene (C) permanently, disabling it.
By itself, it can't move. It's stuck. Ds
is a non-autonomous element.
Third row: Ds and Ac both present, Ds can now excise from
the C gene in some cells (i.e., it can transpose)
during development, creating developmental fields that can produce pigment. This
is because Ac has provided the elements needed for Ds to transpose.
Fourth row: Ac is inserted into pigment gene, but not permanently, as it can
provide the elements that allow its removal from the gene. Ac
is an autonomous element.
Top row: Wild type pigmented kernel.
Second row: Ds is inserted into pigment gene (C) permanently, disabling it.
By itself, it can't move. It's stuck. Ds
is a non-autonomous element.
Third row: Ds and Ac both present, Ds can now excise from
the C gene in some cells (i.e., it can transpose)
during development, creating developmental fields that can produce pigment. This
is because Ac has provided the elements needed for Ds to transpose.
Fourth row: Ac is inserted into pigment gene, but not permanently, as it can
provide the elements that allow its removal from the gene. Ac
is an autonomous element.And the kicker: Rarely, an Ac type was sometimes
found to transform into the Ds type, apparently because the Ac element
spontaneously turned into a Ds element. (This could mean that Ds is simply a
mutant version of Ac that has lost the ability to encode the elements that allow
it to jump around.)
When McClintock first reported her findings in the 1960s, most people
believed that this was something unique to corn. But later, as transposable
elements were discovered in E. coli, yeast, and higher organisms, it became
apparent that she had been the first to describe a phenomenon that was far more
universal, suggesting that genomes were far more dynamic than first supposed. In
1983, she was awared the Nobel Prize in Physiology or Medicine for her early
work on corn transposons.
Several models have been proposed for transposon insertion mechanism have
been proposed. The simplest and most elegant may be that of J.
Shapiro. It partly explains the presence of direct and/or inverted repeats
where transposons insert.
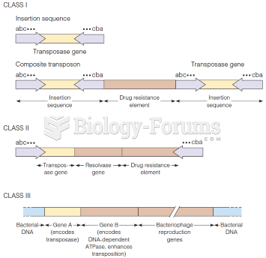
Insertion Sequences: Prokaryotic Transposable Elements Insertion
sequences (IS) were first discovered
in the gal operon
of E. coli, and were
physically located because viruses carrying the bacterial gene in both mutated
and wild type forms could be separated in a centrifuge: the mutants had an extra
piece of DNA inserted, making them denser.
When an IS appears in any of the three genes of the gal operon (E for
epimerase, T for transferase and K for kinase), the normal transcription of the
gene is disrupted.
Insertion of an IS affects only the transcription of the genes downstream from
the insertion. For example, if the IS occurs late in the E gene, the T and K
genes might be disrupted, but the E might not be, and epimerase is still
manufactured.
This phenomenon is known as a polar
mutation, since there is directionality to the transcriptional effects.
Transposons: More Prokaryotic Mischief
In the 1950's a strain of Shigella bacteria
appeared in Japanese hospitals. The normal strains of this bacterium are
sensitive to a wide spectrum of antibiotics. But a Shigella strain isolated from
patients with a severe dysentery, was discovered to be resistant to most
antibiotics.
The multiple-drug resistance phenotype was apparently inherited as a single
package--and not only by other Shigella.
Other bacterial species could also obtain this resistance.
The problem was a self-replicating
episome, a bacterial genetic element capable of
- replicating freely in the cytoplasm or
- being inserted into the bacterial chromosome to replicate along with the
chromosome
This episome was called an R factor (for
"Resistance").The R factor is transferred rapidly between bacteria upon
conjugation. In the cytoplasm, it exists as a plasmid.
As you may recall, plasmids in bacteria often carry genes that confer
resistance to antibiotics
If one denatures the DNA of these R Factors and allows them to slowly
renature, portions of the plasmid form a stem
loop.
The genes conferring drug resistance are usually located on the LOOP of the
stem loop. This is located between two inverted
repeat (IR) sequences, which create the stem loop.
The resistance genes in the loop, along with their flanking IR sequences are
known as a transposon. The regions
between the IR sections are known as the resistance
transfer region (RTR), since that's what carries the antibiotic resistance
genes.
A transposon can jump from one plasmid to another, or directly into the
bacterial chromosome.Two mechanisms
for transposition are known in
prokaryotes:
Both mechanisms generate
a repeated sequence of the target
DNA (i.e., the DNA in which the transposon is inserted).
Although transposons may excise without affecting surrounding DNA, they often
generate a high incidence of deletions in their vicinity. These can consist of
part of the element and part of the adjacent DNA.
When varying lengths of the surrounding DNA are excised along with the
transposon, imprecise excision is
said to have taken place.
When the transposon is excised and deleted portions of the adjacent DNA are
restored, precise excision is
said to have taken place.
Imprecise excision is far more common than precise excision.
Phage µ
This temperate virus (a bacteriophage) inserts into the genome of E.
coli.
If more than one µ is present, they can cause deletions, insertions and
translocations of the host's chromosome if both excise at once.
µ replicates with the host c'some, and generally does not form a plasmid.
Transposable Genetic Elements in Other Eukaryotes Transposable
Genetic Elements have also been found in yeast. Among them are
- retrotransposon -
transposable element in yeast that creates an RNA intermediate (via the
activity of reverse transcriptase) to effect transposition (It acts a lot
like a retrovirus which--when copied into double-stranded DNA and inserted
into the host genome--is called a provirus.)
Ty elements - found in yeast, hese
are characterized by direct repeats at each end.As much as 10% of Drosophila's genome
may consist of families of dispersed,
repetitive DNA sequences that move about as discrete units. Three general
types are known and named:
Let's have a closer look at P Elements P
Elements were first discovered due to
a phenomenon--observed in controlled laboratory matings--known as hybrid
dysgenesis (a fancy term for "many
things wrong with the hybrid") in offspring produced in a cross of M (maternal)
cytotype females (known in the lab only) and P (paternal/wild type) cytotype
males. Problems included sterility, and appearance of weird mutations.
The Matings:
Why?

- A large percentage of dysgenic flies (sterile at high temperatures, but
reproductive at normal temperatures) showed evidence that the dysgenic
mutations could be easily and frequently reversed,
usually in the germline.
- The flies with very high reversion to wild type are generally those with
the M cytotype.
- Hypothesis: the mutations are being caused by the insertion of foreign
DNA--which could later readily and spontaneously excise, reversing the
mutations.
- Probing (with an eye color gene) to recover a particular dysgenic gene
revealed that most of the white eye mutations in dysgenic offspring were
caused by the insertion of a genetic element into the wild type eye color
locus (white eyes resulted from disruption of the wild type red pigment
deposition gene).
- This element was named the P
element.
- It is present (30-50 copies per genome) in P cytotype flies, but completely
absent in M cytotype flies
- The P element can be 0.5 - 2.9 kb in length, but it's always flanked by
a 31-base, perfect inverted repeat (red flag for transposable element)
- The complete P element encodes four genes, one of them an enzyme coding
for a transposases (what
do you suppose this enzyme facilitates?).
- The current hypothesis: The P element encodes not only transposase, but
also repressor products that inactivate
transposase.
- In P cytotype (wild type) flies, there are many P elements. Thus leads a
high concentration of the P element products are produced in P cytotype
flies.
- Because there are transposase repressors produced along with the
transposase, P elements are immobile: transposase cannot operate on them. No
dysgenic mutations occur.
- Because M cytotype (mutant) flies lack the P element, they also they
lack the repressor protein products that normally are deposited in the
cytoplasm.
- Hence, if the cross of a P cytotype male fly with an M cytotype female
fly will yield an embryo with (1) maternal cytoplasm lacking P elements plus (2)
a male-donated nucleus which has some inserted
P elements.
- In such a hybrid, the P elements will make enough transposase to cause
the P element to jump around and cause dysgenesis, but since no repressors
have been laid down in the M type maternal cytoplasm, the transposase works
fine, causing transposition.
- Apart from being just plain interesting, P elements have become a major
tool for the geneticist working on Drosophila genetics,
since they are very useful for tagging genes to be cloned and for inserting
genes transgenically.
Modern Transposable Genetic Vocabulary
target gene - the gene that is
inactivated by insertion of a transposable genetic element (t.g.e.)
receptor element - the t.g.e.
that's inserted into a gene, inactivating it.
regulator gene - a gene at a
completely separate locus, the product of which causes the receptor element to
move about.
controlling elements - another
name for the receptor element and the regulator gene that makes it move around.
The disrupted gene is said to be nonautonomous.
Its expression depends not only on its own existence, but also on the action of
the controlling elements (and the presence/absence of the regulator).
A gene that is always potentially turning on and off via the insertion of a
receptor element (unpredictable though it might be) is said to be autonomous.
In such a gene, it is probable that the regulator gene (Ac, for example) has
actually inserted itself into the target gene and does the inactivation (by
jumping in) or activation (by jumping out) all by itself.
A World of Eukaryotic TGEs As investigators search across species, it is
becoming apparent that large genomes have tremendous numbers of transposable
elements, and may even be composed mostly of
transposable elements.
This may help explain the C-value
paradox: There appears to be little correlation between the size of an
organism's genome and its biological complexity.
Nearly half of the human genome appears to consist of transposable elements,
mostly long interspersed elements
(LINEs) and short
interspersed elements (SINEs). Most of these can no longer move about, but
retain the vestiges of former mobility (e.g., inverted repeats). A vast number
also are included only in introns, and are excised and never transcribed. They
are evolutionary relics rendered harmless by the points of their insertion and
by the host's regulatory mechanisms.
A few elements, however, are still able to move around, and some are known to
be responsible for causing human disorders by inserting into specific locations:
hemophila B (insertion in the factor IX gene)
neurofibromatosis (insertion in the NF1 gene
one type of breast cancer (insertion in the BRCA2 gene)
This is likely to be only small, initial list. More are undoubtedly going to be
found.
In grasses used by humans for grain production, differences in genome size
can largely be attributed to different quantities of inserted LTR
transposons. Except for the transposon regions, the different grasses show a
great deal of synteny in their genomes.

How is such a massive load of transposons tolerated?
Like any good parasite, a smart transposon doesn't harm its host. The ones that
persist are those that have landed in genetic safe
havens: areas of the genome where there are few functional genes. The
transposons just hang out and are replicated--the ultimate freeloading
passengers.
some elements can be used as biotechnology tools for cloning and gene
manipulation, facilitating insertion of genes into germ lines of recipient
cells.
The properties of TGEs may allow their use in gene therapy: insertion of
functional genes in individuals lacking a normal, functioning copy of an
essential gene (e.g., those with "bubble boy syndrome", who lack the precursor
cells necessary to manufacture important cells of the immune system).So maybe
in the long run, we'll be glad of our little passengers, and they'll eventually
be paying their way by means we can't yet foresee.

 Quick Reply
Quick Reply