|
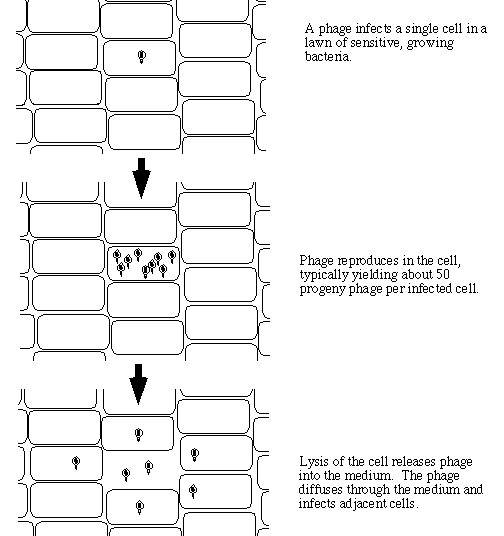
Bacteriophage growth may be differentiated into sequential steps: (i) phage collision with an adsorption-susceptible bacterium, (ii) virion attachment, (iii) virion nucleic acid uptake, (iv) an eclipse period during which infections synthesize phage proteins and nucleic acid, (v) a "post-eclipse" period during which virions mature, (vi) a virion release step, and (vii) a diffusion-delimited period of virion extracellular search for bacteria to adsorb. The latent period begins at the point of virion attachment (ii) and/or nucleic acid uptake (iii) and ends with infection termination, spanning both the eclipse (iv) and the post-eclipse maturation (v) periods.
For lytic phages, latent-period termination occurs at lysis, i.e., at the point of phage-progeny release (vi). A second compound step is phage adsorption, which, depending upon one's perspective, can begin with virion release (vi), may include the virion extracellular search (vii), certainly involves virion collision with (i) and then attachment to (ii) a bacterium, and ends either with irreversible virion attachment to bacteria (ii) or with phage nucleic acid uptake into cytoplasm (iii). Thus, the phage life cycle, particularly for virulent phages, consists of an adsorption period, virion attachment/nucleic acid uptake, a latent period, and virion release. The duration of these steps together define the phage generation time and help to define rates of phage population growth. Also controlling rates of phage population growth is the number of phage progeny produced per infection: the phage burst size. In this chapter we present protocols for determining phage growth parameters, particularly phage rate of adsorption, latent period, eclipse period, and burst size.
Binary fission begins with DNA replication. DNA replication starts from an origin of replication, which opens up into a replication bubble (note: prokaryotic DNA replication usually has only 1 origin of replication, whereas eukaryotes have multiple origins of replication). The replication bubble separates the DNA double strand, each strand acts as template for synthesis of a daughter strand by semiconservative replication, until the entire prokaryotic DNA is duplicated.
Each circular DNA strand then attaches to the cell membrane. The cell elongates, causing the DNA to separate.
Cell division in bacteria is controlled by the FtsZ, a collection of about a dozen proteins that collect around the site of division. There, they direct assembly of the division septum. The cell wall and plasma membrane starts growing transversely from near the middle of the dividing cell. This separates the parent cell into two nearly equal daughter cells, each having a nuclear body. The cell membrane then invaginates (grows inwards) and splits the cell into two daughter cells, separated by a newly grown cell plate.
In summary:
1- the cell grows
2- the DNA is copid and attached to the cell membrane
3- the DNA and its copy separate as the cell rapidly grows larger
4- the cell splits in 2; each new cell has a identical copy of the DNA
|
 Quick Reply
Quick Reply