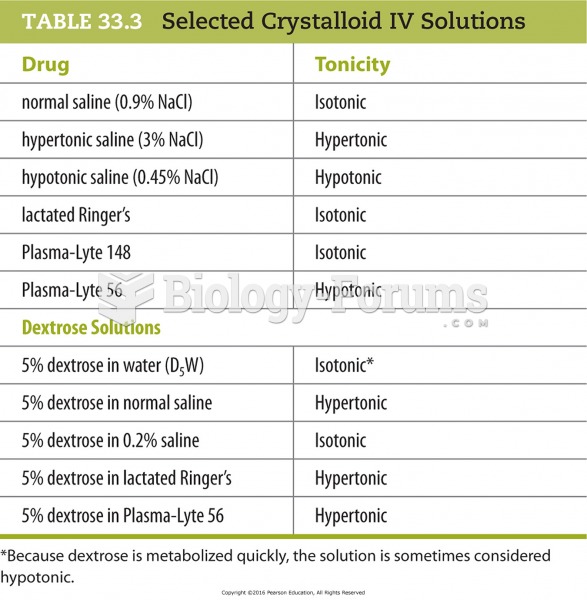
P. 529, #2. A variety of squash has fruits that weigh about 5 pounds each. In a second variety, the average weight is 2 pounds. When the two varieties are crossed, the F1 produce fruit with an average weight of 3.5 pounds. When two of these are crossed, a range of fruit weights is found, from 2 to 5 pounds. Of two hundred offspring, three produce fruits weighing 5 pounds and three produce fruits 2 pounds in weight. How many allelic pairs are involved in the weight difference between the varieties, and approximately how much does each effective gene contribute to the difference?To determine the number of loci that affect the trait, we need to remember the relationship that gives the proportion of homozygous recessive individuals in the F2 generation for any n-hybrid cross. In the case of quantitative traits this proportion is the frequency with which we expect the extreme phenotypes to occur. This proportion is
((1/2^n))^2
where n is the number of loci involved. In this case we set this proportion to the frequency of occurrence of one of the extreme phenotypes, say the 2 pound variety in the F2.
3/200 = ((1/2^n))^2 and solve for n
0.015 = ((1/2^n))^2 then take the square root of each side
0.1225 = (1/2^n) then take reciprocal of both sides
8.165 = 2^n then determine the closest integral value of n that approximates 8.
3 = n thus, there are three loci that affect fruit weight in squash.
To determine how much each effective allele adds to weight, we take the phenotypic value of one parent and subtract from it the phenotypic value of the other parent, then divide that number by twice the number of loci, which is the total number of effective alleles possible.
phenotypic value of one parent - phenotypic value of other parent
= ----------------------------------------------------------------
2 * (number of loci)
5 - 2
= -------
2 * 3
= 0.5 lb per allele
P. 529, #3. In rabbit variety 1, ear length averages 4 inches. In a second variety, it is 2 inches. Hybrids between the varieties average 3 inches in ear length. When these hybrids are crossed to each other, the offspring exhibit a much greater variation in ear length, ranging from 2 to 4 inches. Of five hundred F2 animals, two have ears about 4 inches long and two have ears about 2 inches long. Approximately how many allelic pairs are involved in determining ear length, and how much does each effective gene seem to contribute to the length of the ear? What do the distributions of P1, F1, and F2 probably look like?To determine the number of loci that affect the trait, we need to remember the relationship that gives the proportion of homozygous recessive individuals in the F2 generation for any n-hybrid cross. In the case of quantitative traits this proportion is the frequency with which we expect the extreme phenotypes to occur.
This proportion is
((1/2^n))^2
where n is the number of loci involved. In this case we set this proportion to the frequency of occurrence of one of the extreme phenotypes, say the 2 inch variety in the F2.
2/500 = ((1/2^n))^2 and solve for n
0.004 = ((1/2^n))^2 then take the square root of each side
0.0625 = (1/2^n) then take reciprocal of both sides
15.8 = 2^n then determine the closest integral value of n that approximates 16.
4 = n thus, there are three loci (allelic pairs) that affect ear length in rabbits.
To determine how much each effective allele adds to ear length, we take the phenotypic value of one parent and subtract from it the phenotypic value of the other parent, then divide that number by twice the number of loci, which is the total number of effective alleles.
phenotypic value of one parent - phenotypic value of other parent
= ----------------------------------------------------------------
2 * (number of loci)
4 - 2
= -------
2 * 4
= 0.25 inches per allele
The P1 and F1 distributions are quite narrow (low variance)because all individuals have almost the same genetic composition and thus there is little variability among the individuals. On the other hand, the distribution of the F2s are quite broad. This is because the individuals have different genotypes and thus are quite variable in their phenotypic expression. The variance is much higher in the F2 generation than in the P1 and F1 generations.
P. 529, #4. Assume that height in people depends on four pairs of alleles. How can two persons of moderate height produce children who are much taller than they are? Assume that the environment is exerting a negligible effect.A moderate height person will have genotype with 4 contributing alleles among 4 loci and that person could be heterozygous for contributing and noncontributing alleles at each locus (AaBbCcDd). If that person mates with another person who also has a similar genotype, then the offspring could inherit mostly (or all) contributing alleles from each parent. The offspring could have more contributing alleles than either parent and thus be taller than either parent.
P. 530, #8. Would you expect good nutrition to increase or decrease the heritability of height?Good nutrition would increase the heritability of height because it would reduce the phenotypic variance in the population by reducing the variance due to the environment. Remember that heritability in the narrow sense (hn^2) = Va/Vp. By reducing Vp and leaving Va unchanged, the heritability is increased.
P. 530, #10. If skin color is caused by additive genes, can marriages between individuals with intermediate-colored skin produce light-skinned offspring? Can such marriages produce dark-skinned offspring? Can marriages between individuals with light skin produce dark-skinned offspring?This is similar to problem #4 above, but instead of height the trait is skin color. A person with intermediate-colored skin will have a genotype with an intermediate number of contributing alleles among the loci and that person could be heterozygous for contributing and noncontributing alleles at each locus. If that person mates with another person who also has a similar genotype, then the offspring could inherit mostly (or all) non-contributing alleles from each parent. The offspring could have more non-contributing alleles than either parent and thus be lighter-skinned than either parent.
On the other hand, the offspring could inherit mostly (or all)contributing alleles from each parent. The offspring could have more contributing alleles than either parent and thus be darker-skinned than either parent.
Marriages between light-skinned parents will produce only lighter-skinned offspring. This is because the parents have few to no contributing alleles and thus have few contributing allele to pass to offspring. Thus, the offspring will have few contributing alleles and be light-skinned.
P. 530, #13. The components of variance for two characters of D. melanogaster are shown in the following table. Estimate the dominance and epistatic components and calculate heritabilities in the narrow and broad sense. Variance Thorax Eggs Laid
Components Length In 4 Days
---------------------------------------------------------
Vp 100 100
Va 43 18
VE 51 38
Vd + Vi ? ?
---------------------------------------------------------
If you remember that total phenotypic variance (Vp) is the sum of additive genetic variance (Va) + variance due to environment (VE)+ variance due to dominance (Vd) + variance due to epistasis (Vi), we can use the formula
Vp = Va + Vd + Vi + VE
and by substituting values from the table above into the equation we can solve for the unknown, in this case (Vd + Vi). Thus for thorax length Vd + Vi = 100 - 43 - 51 = 6. We can then use the formulas for heritability in the narrow sense (hn^2 = Va/Vp)and heritability in the broad sense (hb^2 = (Va+Vd+Vi)/Vp) to calculate these uantities.
For thorax length heritability in the narrow sense is
hn^2 = Va/Vp = 43/100 = 0.43
while
heritability in the broad sense is
hb^2 = (Va+Vd+Vi)/Vp = (Va + Vd + Vi)/Vp = 49/100 = 0.49.
For eggs laid in four days Vd + Vi = 100 - 18 - 38 = 44
and
heritability in the narrow sense is
hn^2 = Va/Vp = 18/100 = 0.18 while
heritability in the broad sense is
hb^2 = (Va+Vd+Vi)/Vp = (Va + Vd + Vi)/Vp = 62/100 = 0.62.
P. 530, #14. Psychologists refer to defecation rate in rats as "emotionality". The chart (graph) at the top of page 531 (data modified from Broadhurst. 1960. Experiments in Personality. vol. 1. London: Eysenck) shows mean emotionality scores during five generations in high and low selection lines. In the final generation, the parental mean was 4 for the high line and 0.9 for the low line. The cumulative selection differential is five for each line. Calculate realized heritability overall and separate heritabilities for each line. Do these differ? Why? Why was the response to selection asymmetrical?The response to selection is equal to the realized heritability times the selection differential. In this case, we are given the selection differential (5 for each line) and the response to selection (4 - 3 (1) for the high line and 3 - 0.9 (2.1) for the low line, and are asked to calculate the realized heritability (hn^2). Thus, realized heritability is equal to the response to selection (R) divided by the selection differential (S).
hn^2 = R/S
The overall realized heritability will combine both lines. The selection differential (S) = 5 + 5 = 10 and the response to selection (R) = 4 - 0.9 = 3.1.
hn^2 = R/S = 3.1/10 = 0.31
The realized heritability for the low line will use just the lower part of the graph. The selection differential (S) = 5 and the response to selection (R) = 3 - 0.9 = 2.1.
hn^2 = R/S = 2.1/5 = 0.42
The realized heritability for the high line will use just the upper part of the graph. The selection differential (S) = 5 and the response to selection (R) = 4 - 3 = 1.
hn^2 = R/S = 1/5 = 0.20
The realized heritability for the two lines is different. This is probably due to the frequency of contributing alleles in the original population. If the frequency of contributing alleles was fairly high then you would not expect selection to increase their frequency much. On the hand, if the contributing alleles are less common, then selection can go a long ways in increasing the frequency of the non-contributing alleles (and thus a greater response to selection.
P. 531, #18. A plant with a genotype of aabb and a height of 40 cm is crossed with a plant with a genotype of AABB and a height of 60 cm. If each dominant allele contributes to height additively, what is the expected height of the F1 progeny?In this case, as the parents are both homozygous, the F1 progeny will be heterozygous at both loci and thus have half the number of dominant alleles that the parent that is homozygous for dominant alleles has. This means that the F1 offspring will be intermediate in height between the two parental genotypes. Thus the height of the F1 is simply an average of the parental heights. (60 + 40)/2 = 50 cm.
To determine how much each dominant allele adds to height, we take the phenotypic value of one parent and subtract from it the phenotypic value of the other parent, then divide that number by twice the number of loci, which is the total number of dominant alleles.
phenotypic value of one parent - phenotypic value of other parent
= ----------------------------------------------------------------
2 * (number of loci)
60 - 40
= ---------
2 * 2
= 5 cm per allele
P. 531, #20 You determine the following variance components for leaf width in a particular species of plant:
Additive genetic variance (Va) 4.0
Dominance genetic variance (Vd) 1.8
Epistatic variance (Vi) 0.5
Environmental variance (Ve) 2.5
Total phenotypic variance (Vp) is the sum of additive genetic variance (Va) + variance due to environment (VE) + variance due to dominance (Vd) + variance due to epistasis (Vi). Thus, we can use the formula
Vp = Va + Vd + Vi + VE
to calculate Vp, which we were not given.
Vp = 4.0 + 1.8 + 0.5 + 2.5 = 8.8 To calculate the broad sense and narrow sense heritability, we can use the formulas for heritability in the narrow sense (hn^2 = Va/Vp) and heritability in the broad sense (hb^2 = (Va+Vd+Vi)/Vp). For leaf width heritability in the narrow sense is
hn^2 = Va/Vp = 4.0/8.8 = 0.45
while
heritability in the broad sense is
hb^2 = (Va+Vd+Vi)/Vp = (Va + Vd + Vi)/Vp = 6.3/8.8 = 0.72.
P. 531, #22 The narrow sense heritability of egg weight in chickens in one coop is 0.5. A farmer selects for heavier eggs by breeding a few chickens with heavier eggs. He finds a difference in the mean egg weight of 9 g between selected and unselected chickens. By how much can he expect egg weight to increase in the selected chickens?Remember that response to selection (R) is a function of heritability in the narrow sense (hn^2) and the selection differential (S). The relationship is given by the following formula
R = (hn^2)*S
In the case of the farmer, the difference between selected and unselected chickens for egg weight (9 g) is the selection differential and heritability in the narrow sense is 0.5. We can use the above formula to calculate the response to selection, which is how much the average egg weight should increase in the offspring of selected chickens.
R = (hn^2)*S = 0.5*9 = 4.5 grams
Thus the response to selection is 4.5 grams.
P. 531, #24 Two strains of wheat were compared for the time required to mature. Strain X required 14 days and strain Y required 28 days. The strains were crossed, and the F1 generation was selfed. One hundred F2 progeny out of 6,200,000 matured in 14 days or less. How many genes may be involved in maturation?
To determine the number of loci that affect the trait, we need to remember the relationship that gives the proportion of homozygous recessive individuals in the F2 generation for any n-hybrid cross. In the case of quantitative traits this proportion is the frequency with which we expect the extreme phenotypes to occur.
This proportion is
((1/2^n))^2 where n is the number of loci involved.
We set this proportion equal to the frequency of occurrence of one of the extreme phenotypes, in this case, the strain that matures in 14 days or less in the F2.
100/6,200,000 = ((1/2^n))^2 and solve for n
0.000016 = ((1/2^n))^2 then take the square root of each side
0.004 = (1/2^n) then take reciprocal of both sides
248.99 = 2^n then determine the closest integral value of n that approximates 256. You need to keep in mind the powers of 2.
8 = n thus, there are eight loci (allelic pairs) that affect maturation time in wheat.
P. 531, #25. If, in a population of swine, the narrow sense heritability of maturation weight is 0.5, the phenotypic variance is 100 lb2, the total genetic variance is 50 lb2, and the epistatic variance is 0, calculate the dominance genetic variance and the environmental variance. Remember
Vp = Va + Vd + Vi + VE
and genetic variance (VG) = Va + Vd + Vi and heritability in the narrow sense (hn^2) = Va/Vp. By knowing that hn^2 = 0.50, we can determine that Va = 50.
Va = Vp*hn^2 = 100 * 0.50 = 50
By knowing that VG = 0.50 and Vi = 0, we can determine that Vd = 0.
Vd = VG - Va - Vi = 50 - 50 - 0 = 0
By knowing the VG = 50 and Vp = 100, we can determine that VE = 50.
VE = Vp - VG = 100 - 50 = 50.
P. 531, #26. A group of four-month-old hogs has an average weigth of 70 pounds. The average weight of selected breeders is 185 pounds. If the heritability of weight is 40%, what is the expected average weight of the first generation progeny?Remember that response to selection (R) is a function of heritability in the narrow sense (hn^2) and the selection differential (S). The relationship is given by the following formula
R = (hn^2)*S
In the case of the farmer, the difference between selected (185 pounds) and unselected hogs (170 pounds) for weight is the selection differential (15 pounds) and heritability in the narrow sense is 0.40 (40%). We can use the above formula to calculate the response to selection, which is how much the average weight should increase in the offspring of selected hogs.
R = (hn^2)*S = 0.4*15 = 6.0 pounds
Thus the response to selection is 6.0 pounds. You then add this amount to the average weight before selection (170 pounds). The average weight of the progeny after selection is 176 pounds.

 Quick Reply
Quick Reply