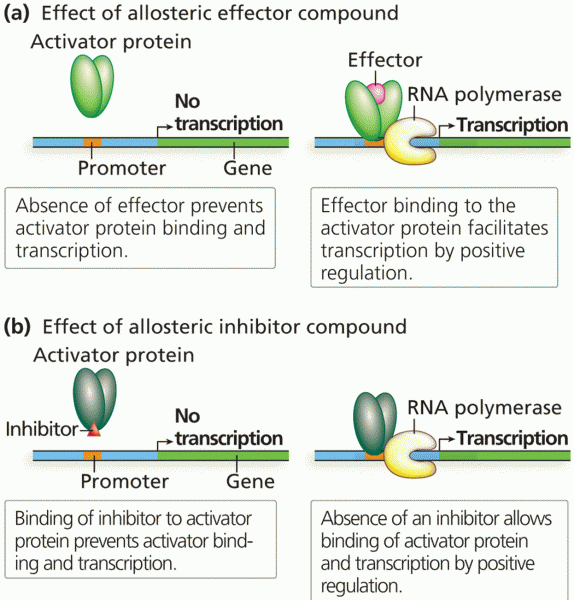
To be more specific: Yes, you will have to include what you said: (effectors , modulators Rf orm and T from MM kinetics and sigmoid curves - remember that sigmoid curves are used instead of MM curves at allosteric enzymes)
You will also have to write examples of the advantages of allosteric regulation.
An obvious example is hemoglobin. As you know, hemoglobin is consisted by four polypeptide chains, and each chain can bind with on oxygen or CO2 molecule.
If one (or more) of the chains are binded to oxygen, then, an allosteric change will increase hemoglobin's affinity to oxygen.
Respectively, if one (or more chains) are binded with CO2 (and H+), then the hemoglobin's affinity to CO2 will increase,
and the affinity to O2 will decrease.
The advantage of this example are:
When hemoglobin is found to an environment with high concentration of oxygen, then much more oxygen will bind to hemoglobin.
When hemoglobin is found to an enfironment with low oxygen and (relatively) high concentration of CO2, then much more O2 will be released, and much more CO2 will be binded (compared to the case of hemoglobin not being allosteric)
From stryer's biochemistry:
Hemoglobin Transports Oxygen Efficiently by Binding Oxygen Cooperatively
Hemoglobin, the oxygen carrier in the blood, is an allosteric protein. Hemoglobin consists of four polypeptide chains, each with a heme group a substituted porphyrin with a central iron. Hemoglobin A, the predominant hemoglobin in adults, has the subunit structure α2 β2. Hemoglobin transports H+ and CO2 in addition to O2. Hemoglobin exhibits three kinds of allosteric effects. First, the oxygen-binding curve of hemoglobin is sigmoidal, which indicates that the binding of oxygen is cooperative. The binding of oxygen to one heme group facilitates the binding of oxygen to the other heme groups in the same molecule. Second, the binding of H+ and CO2 promotes the release of O2 from hemoglobin, an effect that is physiologically important in enhancing the release of O2 in metabolically active tissues such as muscle. These allosteric linkages between the binding of H+, CO2, and O2 are known as the Bohr effect. Third, the affinity of hemoglobin for O2 is further regulated by 2,3-bisphosphoglycerate (2,3-BPG), a small molecule with a very high density of negative charge. 2,3-Bisphosphoglycerate binds tightly to deoxyhemoglobin but not to oxyhemoglobin. Hence, 2,3- BPG lowers the oxygen affinity of hemoglobin. Fetal hemoglobin (α2 γ2) has a higher oxygen affinity than human adult hemoglobin because fetal hemoglobin binds 2,3-BPG less tightly. Neither the sequential nor the concerted model completely describes the allosteric behavior of hemoglobin. Rather, the behavior of hemoglobin is best described by a combined model that employs features of both models.
Generally, allosteric regulation provides better regulation of enzyme reaction and protein's use.
 Quick Reply
Quick Reply